Mi a baj az evolúcióval? (javítva)
2010. 01. 19.
Korábban emlegettük már az evolúció-elmélet helytelen, hiányos és pontatlan, csalóka voltát, itt az ideje, hogy végre tisztázzuk: miért nem fogadjuk el a darwinizmust?
I. Az evolúció története
Az evolúcióelmélet alapjait a közhiedelemmel ellentétben nem Charlie Darwin rakta le. Már az ókorban is léteztek „szinogenetikus” elképzelések, tehát a különböző típusú élőlények közös származásáról szóló filozófiai tételek. Természetesen ezek nemigazán „genetikai” származást vagy rokonságot jelentenek, hiszen a modern genetika alapjait csak Mendel rakta le a 1865-ben. Sokkal inkább az „asztrális szimpátia” elvére épültek, amely a természet okkultista szemmel történő tanulmányozásából fakadt. (Az ókorban és a kora középkorban minden tudós okkult beavatott is volt.)
Az evolúció és a közös eredet modern megközelítése, amelyet darwinizmusnak is hívnak, – nagyon meglepő módon – egy titkos társaságtól eredeztethető, amelyet Lunáris Körnek neveztek. Ennek tagja volt Erasmus Darwin, Charlie nagypapája, aki 1796-ban elsőnek fogalmazta meg részletesen az evolúció menetének mai elképzelését, faszerkezet-jellegű hierarchiát tulajdonítva az élőlények típusainak, s egyetlen közös szó szerinti őst feltételezett. Nézeteit a korabeli (akkor még keresztény) tudósok nagy része nem fogadta el. A társaságnak mellesleg szintén tagja volt Benjamin Franklin, valamint – ami a lényeg – egy James Hutton nevű skót polihisztor, akinek a nevét nem különösebben jegyezte fel az utókor (legalábbis a tankönyvekből valahogy rendre kimarad szegény), pedig ő állt elő az ötlettel, hogy a Föld több millió éves (akkor még csak annyi volt; Kent Hovind kiszámolta, hogy a tudósok percenként 41 évet öregítenek bolygónkon). Elmélete, a „plutonizmus” felváltotta a korabeli „neptunizmust” (amely alátámasztotta a fiatal Föld elképzelést s a vízözönről szóló, minden vallásban jelen lévő történetet), megosztotta a tudomány világát, s utat nyitott Charles Lyellnek.
Charles Lyell egy skót jogász volt, tudományos végzettsége nem volt, bár végigülte William Buckland néhány ásványtan óráját 1816-ban, majd 1821-ben Robert Jameson szemináriumát (mindkét tudós Hutton lelkes híve volt), s ez „különös módon” feljogosította arra, hogy a Londoni Geológiai Társaság titkárává váljon 1823-ban. 1830-ban már geológia professzor volt a Londoni Királyi Főiskolán (lehet, hogy az intézménynevekre és könyvcímekre nem az általánosan elfogadott fordítást használom, de angol szakirodalomból dolgozok, a magyar szokás szerint sovány, pontatlan – marxista újradoktrinálás – és nehezen hozzáférhető), és megjelentette a Geológia Alapelvei című könyvét, amely mérföldkő volt mind a geológia, mind az evolúciós biológia történetében. Ebben pontosan meghatározta az egyes geológiai rétegek korát, „bebizonyítva”, hogy a Föld – immár – több százmillió éves, s az egyes rétegek egyes földtörténeti korokból származnak, ahogy azt ma a világon minden iskolában tanítják. A különböző rétegekben pedig különböző fejlettségű állati fosszíliák találhatóak, s ez geológiai bizonyíték: az evolúcióra.
Hogy mindezt Lyell honnan vette, rejtély. 98 évvel járunk azelőtt, hogy a Geiger-Müller számlálót feltalálták volna, a radiometrikus kormeghatározás módszerei pedig csak a XX. század derekán kezdtek megjelenni. Lyell egyszerűen saját korának mérhető geológiai folyamatait (elsősorban a vulkanikus tevékenységet) lineárisan visszaszámolta a múltra, abból kiindulva, hogy több millió (sőt, százmillió) évvel ezelőtt is pontosan ugyanígy zajlottak ezek a folyamatok. A modern geológia ezt az elképzelést réges-régen megdöntötte: emlékezzünk a különböző földtörténeti korok különböző folyamataira, először a röghegységek, majd a lánchegységek jöttek létre; ahogy a földköpeny egyre hidegebb lesz, a vulkanikus tevékenység csökken – minderről Lyell még semmit sem tudhatott! Azonban a geológiai rétegekről szóló meséjét a mai napig nem kérdőjelezte meg a „hivatalos tudomány” – főleg azért, mert a darwinizmus legfőbb „bizonyítékáról” beszélünk.
Lyellnek ugyanis volt egy jóbarátja, akit Charles Darwinnak hívtak. Az ő életútja és munkássága elég közismert és rengeteg anyagot lehet találni róla az Interneten is, tehát erre most ne térjünk ki. A lényeg az, hogy Lyell nézetei és könyve lassan felőrölték előbb a Biblia hitelességébe, majd Istenbe vetett hitét; bár ő maga nem így vallott a nyilvánosság előtt, de a Beagle fedélzetén írott útinaplója erről tanúskodik. Kutatásainak eredményeiben Lyell és saját nagyapja elméleteinek bizonyítékát látta, s elkezdett dolgozni a saját, „pontosított” evolúció-elméletén. Azért az ő nevéhez fűzi a populáris tudomány az evolúciós biológia kezdetét, mert tőle származik a mutáció és természetes szelekció gondolata, amelynek segítségével már pontos magyarázatot tudott adni arra, hogy az egyes fajok végül is hogyan alakultak ki…
S ahogy mindössze 6 évvel később Mendel megerősítette a mutáció létét (vagyis hogy az utódban valóban meg tudnak jelenni olyan genetikai tulajdonságok, amelyek egyik szülő genotípusában sem voltak benne) egyesek azonnal „bizonyítottnak” látták Darwin elméletét – s ez a mai napig is így van.
II. Az evolúció bizonyítékai
Az evolúció egy munkahipotézis. Több mint egy szipla hiedelem, de semmiképpen nem tény. Az evolúció hívei az iskolai diákoknak és általában a nagyközönségnek úgy állítják be, mintha az evolúciónak rengeteg bizonyítéka lenne. Valójában egyetlen olyan bizonyíték sincs, amely például egy bíróság előtt megállná a helyét. Sok minden alátámasztja az evolúciót, ez tény. Ám a literalista kreacionizmust (a Biblia szó szerinti értelmezésére épülő tudatos teremtés elméletét) is rengeteg tudományos kutatási, mérési eredmény alátámasztja. Bizonyítani nem sikerült még egyiket sem, hiszen ha sikerült volna, a vita már eldőlt volna. A kétségbeesett bizonyítási szándék és az óriási erőfeszítések, amelyeket eme két elmélet hívei tettek – néha hamisítástól sem visszariadva – véleményünk szerint megbízható alapot adnak a feltételezésnek, hogy >egyik elmélet sem igaz<. Mindkettőben van igazság – a darwinizmusban is, mint az mindjárt meglátjuk. A darwinizmus melletti tudományos elkötelezettség egyik legfőbb logikus alapja, hogy használható, tehát működik, mint munkahipotézis, eredményeket lehet vele felmutatni. Azonban a kreacionisták is mutattak már fel eredményeket (pl. magasnyomású oxigénkamrák), persze erről nem szabad beszélni.
De lássuk az evolúció főbb bizonyítékait:
1. és legfontosabb: a geológiai bizonyíték. A Földön már 26 olyan helyet találtak, ahol a fosszíliák az egyes rétegekben darwin elméletét alátámasztandó, a kifejlődési sorrendben és Lyellnek is megfelelő korokhoz köthetően találhatóak meg.
2. Az evolúció ma is megfigyelhető, az élőlények átalakulnak, a megváltozó élettér tudományosan dokumentálható speciációt – fajkeletkezést – okoz (amely önmagában egyértelmű cáfolat arra, hogy minden fajt Isten teremtett volna).
3. Az egyes fosszíliák korát radiometrikus kormeghatározással ellenőrizhetjük, számos olyan fosszília és élettelen kőzet is van, amelyek bizonyítják, hogy Charles Lyell pontosan állapította meg a rétegek korát, Darwin elmélete az élőlények fejlődési sorrendjéről és üteméről pedig szintén helyes.
4. Kémiai, szerkezeti, funkcionális, stb. hasonlóságok vannak az egyes fajok között – ezek a rokonsági hidak gyakran még törzsek, sőt országok között is átívelnek: minden élőlény egy vagy több sejtből áll, minden sejt fehérjéi ugyanabból a 20 aminosavból épülnek fel, mindegyikük DNS szerkezete hasonló, stb. Rengeteg a hasonlóság, mely a rokonságot bizonyítja!
5. Jópár élőlényben találhatóak csökevényes szervek vagy kezdemény-szervek.
6. Az embrionális fejlődés stációi az evolúciós fejlődéssel összhangban vannak.
7. Az életnek az őstengerekben, a „prebiotikus őslevesben” való megjelenését kémiai kísérlettel szimulálta több tudós is (pl. a >Zeitgeist-bejegyzésben< emlegetett Carl Sagan), a kísérlet lényegében sikeres volt. Ammóniából, koncentrált elektromos energia felhasználásával, sikerült fehérjeszintézist megindítani.
8. Egyes fajok több kontinensen is jelen vannak, amely csak úgy képzelhető el, hogy egy helyről származnak, s a kontinensek régen egybefüggő szárazföldet alkottak.
III. Problémák az evolúcióval
Lássuk ezeket a „bizonyítékokat” néhány elhallgatott tény fényében:
1. Amiről nem beszélnek: hogy minimum 26.000 olyan hely is található a Földön, ahol NEM a megfelelő sorrendben vannak a fosszíliák, például madarakat találtak az őshüllők alatti rétegekben, trilobitákat találtak emberrel megegyező rétegekben, emberkéz alkotta használati tárgyakat találtak a legmélyebb rétegekben, még a feketekőszénben is, stb.
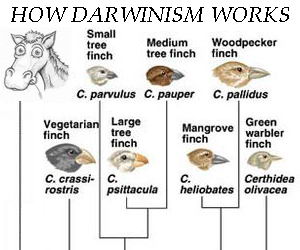
2. Ez a darwinisták legnagyobb csalása: megfogják az egyetlen érvényes és valós részét az elméletnek, és bizonyítékként lobogtatják. Tény: Az élőlények képesek átalakulni. Tény: A természetes kiválasztódás működik. Tény: ha egy adott faj két populációja két különböző élettérre szeparálódik, idővel az eltérő körülmények hatására külön fajjá válnak szét: speciáció. Darwin lényegében a speciációt figyelte meg a Galapagos-szigeteken, különböző madárfajok és különböző hüllőfajok között állapított meg rokonságot – madár a madárral, hüllő a hüllővel. Hogy a hüllő a madárral is rokonságban lenne, meg az árokparti sóskával is, az már csak spekuláció volt a részéről, ennek semmilyen kézzelfogható bizonyítékát nem találta meg (sem senki azóta). Egyszerűen felfedezte, hogy bizonyos fajok rokonságban vannak egymással, s ezt kiterjesztette az egész élővilág összes fajára, részletes és hihető elméletet dolgozott ki arról, hogy az egyes fajok milyen fejlődési sorrendben s milyen fejlődési útvonalon keresztül alakultak ki egymásból. Erről részletesebben a VII. pontban lesz szó.
A speciáció nem cáfolja még a literális kreacionizmust sem, nemhogy tágabb értelmezéseit. A Biblia nem „fajokról” ír (ez a modernkori tudomány kifejezése), hanem fajtákról, típusokról. A kreacionista biológusok nem tagadják a fenti tényeket, mivel azok megfigyelhetőek, mérhetőek, mesterségesen előidézhetőek. Ha mesterségesen változtatott környezetbe helyezünk egy populációt, alkalmazkodni fognak az új viszonyokhoz. Ha zöldre festjük az utcákat, a városi galambok 100 év múlva zöldek lesznek. Ez tény, ezt józan gondolkodású ember nem vitatja – de mivel a kreacionisták sem az akadémikus platformokon, sem a tömegmédiában nem juthatnak szóhoz, ellenfeleik akármit a szájukba adhatnak, s természetesen ezt szokták leggyakrabban a szájukba adni, mivel ez a legnagyobb ostobaság, amellyel legkönnyebben lejárathatóak a tudatos tervezés hívei.
3. Az egyes fosszíliák korát nem radiometrikus kormeghatározással állapítják meg! Az alapján állapítják meg, hogy melyik rétegben találták meg őket. A rétegek korát Charles Lyell adta meg 1830-ban, mindenfajta komolyabb tudományos alap nélkül, s ezeket a számértékeket használják ma is. Ámde az egyes kőzettípusok több rétegben is előfordulnak, így tehát egy adott kőzet adott réteghez és adott földtörténeti korhoz tartozó voltát hogyan állapítják meg? Hogy milyen fosszíliákat találtak bennük! Körkörös érvelés, ahogy a vöröseltolodás kapcsán is említettük a >8. rádióban<; a kormeghatározásnál a természettudomány különféle ágai előszeretettel használnak körkörös érveléseket.
Valóban létezik számtalan olyan fosszília, amelynek kora radiometrikus módszerrel ellenőrizhető, pontos és helyes. Azonban, akárcsak az 1. pont esetében, itt sem beszélnek a milliónyi olyan fosszíliáról, amelyekre hibás értéket ad vissza az adott kormeghatározási módszer. A radiometrikus kormeghatározás egyszerűen nem műdödik. Anomáliás értékeket ad vissza, amelyek közül kiválogatják a tudósok a nekik tetsző értékeket, s bizonyítékként bemutatják a nagyközönségnek. A radiometrikus kormeghatározás talán legmegbízhatóbb módszerének a szénizotópos kormeghatározást tartják, azonban ez egyrészt csak néhány ezer évre visszamenőleg működik pontosan a C14 izotóp viszonylag „rövid” felezési ideje miatt, másrészt valószínűleg erről lesz majd egy külön kiegészítő bejegyzés, példaként demonstrálva, hogy eme „legmegbízhatóbb” módszer hány ponton bukik el.
4. Ezek a hasonlóságok valóban léteznek, azonban ezek nem bizonyítják a közös ős általi rokonságot, csak akként értelmezhetőek az evolúció hívei számára. Éppen ilyen könnyen értelmezhetőek ugyanis a közös tervező általi rokonság jeleként is!
Példa: a rovarok egy része tud repülni, ahogy a madarak nagyrésze is. Nemcsak a repülés képessége közös bennük, de a megvalósítás módja is: a szárnyukat vertikálisan rezegtetik, minél kisebb testűek, annál gyorsabban. Az ember által alkotott repülő szerkezetek egyike sem akalmazza ezt a módszert (héliumos és hőlégballon, sárkányrepülő, légcsavaros és sugárhajtású repülőgép, helikopter, stb.) – az „ornitopter” kizárólag Frank Herbert fantáziájában jelenik meg. Sok módszer van tehát a repülésre, a természet azonban mindkét esetben pontosan ugyanazt használja. Bizonyítja-e ez a közös őst?
Lássuk, létezett-e a darwinizmus szerint olyan „primitív” élőlény a Földön, amely tudott repülni, s mind az ízeltlábú rovarok, mind a gerinces-melegvérű madarak őse lehetett? Természetesen ilyen lény létezését még csak nem is feltételezték soha. Az ízeltlábúak és a gerincesek legközelebbi „közös őse” valahol a lapos- és szalagférgek környékén kereshető talán (majd aki nálam jobban ért hozzá, megmondja). Akkor mit bizonyít inkább ez a közös tulajdonság: közös őst vagy közös tervezőt?
5. Nem léteznek csökevényes szervek A „farokcsont” nem csökevényes szerv, a neve félrevezetés, a csalás része: a gerinces állatok farka a gerincoszlop meghosszabbítása. A „farokcsont” nem része a gerincoszlopnak, funkciója is eltérő (igen, van neki): nem a vázizomzat közvetett tartásában játszik szerepet, hanem a bélcsatorna simaizomszövete* tapad hozzá. „Evolúciósan” tehát semmi köze nincs a farokhoz, ráadásul semmi sem magyarázza, hogy a fáról lejövő s lassan felegyenesedő, emberré váló majomfajtának miért ne lett volna szüksége a farkára továbbra is. Ez propaganda-eszköz, semmi más, a biológusoknak kéne tudniuk a legjobban. A féregnyúlvány sem csökevényes szerv, nagyon fontos része az immunrendszernek, akárcsak az orr- vagy a torokmandula. Ezek nélkül is működőképes marad az immunrendszer, akkor ezek is csökevényes szervek? Fél szemmel is lát az ember, akkor a két szem csökevényes? Nem léteznek funkció nélküli, elcsökevényesedett vagy épp kezdeményszervek – s ha léteznének, az sem bizonyítaná az evolúciót, hanem inkább cáfolná (még annak tényszerű, igaz részét is), hiszen 30-40 generáció alatt eltűnnének.
6. Ezt a hazugságot Ernst Haeckel professzor találta ki 1869-ben. Charlie Darwin lelkes követői ugyanis egy évtized alatt sem tudtak semmilyen bizonyítékot találni az elmélet helyességére. (Nagy dolog. 2009-ben átléptük a kerek 150 évet, és még mindig sehol semmi.) Haeckel tehát úgy döntött, hogy „kreál” egyet! Mindenki emlékszik a biológia-tankönyvből arra az ábrára, amely az egyes állatok és az ember magzati fejlődési stádiumait mutatja be. Ez a rajz minden tankönyvben azonos, mivel Haeckel saját keze munkáját használják mindenhol – ez ugyanis az egyetlen „bizonyíték” az embrionális fejlődés általi evolúciós közösségre. A rajz hamisítvány, amint az egy ultrahanggal ma már könnyedén ellenőrizhető, azonban Haeckel még saját korában is elég hamar lebukott: a Baseli Egyetem professzori kara felfedezte a csalást 1874-ben, s kirúgták az egyetemről. Rajza azonban a mai napig benne van minden tankönyvben (s mellesleg az abortusz-kampány egyik ideológiai sarokköve).
7. Ez egy újabb propaganda-hazugság. (A pontos információkat emlékezetből nyomom, mivel a fellelhető anyag ellentmondásos - mindenhol mást hazudnak.) Problémák a kísérlettel:
a) A kísérlet csak akkor sikeres, ha a rendszerben nincs jelen a bomlasztó hatású UV-sugárzás, s nincs jelen oxigén sem, mivel a szerves anyagok könnyen oxidálódnak. Emiatt találták ki, hogy az élet az óceánok mélyén indult, hiszen ha nincs oxigén, nincs ózonréteg sem, a víz viszont elég jól szűri az UV-sugárzást. A probléma csak az, hogy a víz vezeti az áramot, ezért a víz alatt nincs természetes elektrosztatikusság – mi indította be akkor a folyamatot?
(Apropó, ha az élet a tengerek mélyén indult, ez azt jelenti, hogy 3-4 milliárd évvel ezelőtt már voltak tengerek. Azonban van egy kis probléma: a tengerek folyamatosan elsósulnak, mivel az őket tápláló folyamok hordalékának egy kis része só. A hordalékban lévő por, homok, agyag, stb. leülepszik lassan, a só azonban oldódik a vízben, így addig nem kezd leülepedni, míg az oldat telítetté nem válik. (Ettől még messze vagyunk.) A víz folyamatosan körbejár a párolgás-esőzés által, azonban a só mindig csak egy irányban mozog, nem tud visszakerülni a szárazföldre, így a tengerek egyre sósabbak lesznek. Ha az óceánok több milliárd évesek lennének, már réges-régen telítődtek volna sóval, és az óceánok alatti lerakódások sótartalma is jókora vastagságban megegyezne a százazföldi rétegek sótartalmával. Azonban mivel az oldat telítettségétől még messze vagyunk, az óceáni rétegek sótartalma értelemszerűen sokkal alacsonyabb, közel 0. Vagyis: az óceánok nem lehetnek még egymillió évesek sem…)
b) A végtermék 2%-a volt aminosav. 2% benzol és 96% kátrány keletkezett még. A probléma tehát: a produktum 98%-a mérgező volt az életre, mivel az aminosavak sokkal gyorsabban kötődnek hozzájuk, mint egymáshoz. A kísérlet tehát lényegében kudarc volt.
c) A húszféle aminosavból mindösszesen néhányat, az egyszerűbbeket sikerült előállítani. A molekulák fele „balkezes”, másik fele „jobbkezes” volt, a természetben ez nem így működik, az aminosavak és ribonukleinsavak egynemű bal- és jobbkezességük révén tudnak egymáshoz kapcsolódni, egymásra oda-visssza hatni.
d) Még ha sikeres is lett volna a kísérlet, néhány aminosav nem jelenti azt, hogy „életet” csináltunk. Ha az aminosavakat úgy fogjuk fel, mint az ábécé betűit, akkor a fehérjék a mondatok. Egy sejt legalább egy könyvespolcnyi mondatból áll, s egyetlen betű rossz helyre kerülésétől működésképtelenné válhat. Ha valaki betűtésztából kiszórja nekem a Gyűrűk urát (s az még csak egyetlen kötet a polcról, mondjuk egy kis sejtszervecske), akkor elhiszem, hogy az élet így keletkezett. Ez az elképzelés még a XIX. századból származik, amikor azt hitték, egy sejt az a testnek egy „építőköve”, kis darab zselé, vékony hártyával körülvéve – a korabeli mikroszkópokkal ugyanis ennyit láttak belőle. Ma már tudjuk, hogy egy sejt bonyolultabb, mint bármely emberkéz alkotta szerkezet.
8. A fajok interkontinentális szétszórtsága megmagyarázható azzal is, hogy volt egy vízözön, és előtte nem voltak óceánok.
A kontinensek vándorlása pedig egy bugyuta mese. A kőzetlemezek valóban el tudnak mozdulni, de csak néhány métert. A Föld ugyanis nem madártej, a kontinensek nem az óceánokon lebegnek, éppen ellenkezőleg: az óceánok jelentéktelen tömegű és vastagságú pocsolyák a kontinentális lemezekhez képest. Az óceán felszíne alatt mindössze néhány kilométerrel ugyanolyan kőzetlemez található, mint amelyen most ülünk. Ha a kontinensek ezer kilométereket mozognának, sok száz kilométeres lánchegységek gyűrődnének fel. Ezek hol vannak?
A kontinensek összeilleszthetőek, ezt láthatjuk a tankönyvek ábrázolásain. Ezzel két probléma van:
a) Nem a kontinensek alakja a döntő, hanem a kontinentális lemezek alakja, az pedig nem összeilleszthető. A kontinensek alakja a vízszint függvényében változik, a vízszint pedig a sarki jég olvadása miatt folyamatosan emelkedik (az emberi tevékenység csak meggyorsította ezt a folyamatot), vagyis a kontinensek X millió évvel ezelőtt teljesen más alakúak voltak.
b) A kontinensek nem összeilleszthetőek, a tankönyvekben látható rajz hamisítvány, ez könnyen ellenőrizhető egy műholdas térkép segítségével: a rajzon a kontinensek nem méretarányosak. Afrika és Dél-Amerika arányában van a legnagyobb eltérés, erről bárki könnyen meggyőződhet.
IV. A Darwinizmus halála
Darwin szerint az evolúció receptje: véletlen mutáció + természetes szelekció. Ennek mindkét fele megdőlt mostanra.
Lássuk a természetes szelekciót! Ennek lényege, hogy az adott kontextusban – élőhely/környezet/társulások/verseny/stb – a nagyobb eséllyel túlélő és szaporodó egyed az életképesebb, s az ő génjei nagyobb arányban keverednek be a populációba. De hogy nem minden faj ezen folyamat eredménye, azt már Darwin korában is jól tudták az emberek. Nézzük a tehenet! A tej „evolúciós” szerepe, hogy az újonnan születő utódnak átmeneti, de teljes értékű táplálékot biztosítson – az anya anyagcseréjéről az utód szülés után, fokozatosan válik le. A tehenek azonban nemcsak olyankor adnak tejet, mikor borjuk van, hanem folyamatosan. Ez egy mutáció eredménye. Mitől hasznos ez a mutáció? Nekünk abban, hogy megisszuk a tejet, a tehénnek legfeljebb annyiban, hogy mi döntöttük el a múltban, melyik tehenet engedjük szaporodni, tehát ha több tejet ad, nagyobb az esélye a túlélésre és szaporodásra. Amit tehát itt látunk, az mesterséges szelekció, nem természetes!
A véletlen mutáció szintén megdőlt a ’80-as években, s itt újra, sokadszorra hivatkoznék Bruce Lipton előadására – bár aki eddig nem nézte meg, most se fogja. Az élőlények DNS-e nem egy könyvhöz, inkább egy füzethez hasonlít, amely íródik is az élettapasztalatok függvényében. Az írás módját és mikéntjét szintén erre a célra létező, „génmodósító gének” határozzák meg – persze nem maga a gén végzi a módosítást, a gének közvetlenül nem csinálnak semmit, a fent említett balkezes-jobbkezes minta alapján a ribonukleinsavak (ideértve a dezoxi-ribonukleinsavat is) az aminosavakra hatnak, s vice versa. A gének önmagukban nem „csinálnak” semmit, passzív elemek, fizikai tevékenységet csak az aminosavak, ill. a belőlük felépülő fehérjék végeznek. A „DNS redundanciája” egy baromság, amely vissza-visszatér a genetikai kutatásokról szóló cikkekben: a gének nagyrésze ugyanis olyan viselkedésformák receptjeit tartalmazza, amelyekre a fehérjéknek egy adott, monoton élettérben nincs szüksége, így ezek az információk nem hívódnak le. De amint az élettér megváltozik, le fognak hívódni!
A gének és az élőhely tehát folyamatos egymásrahatásban vannak az élőlényeken keresztül, random mutációk pedig, bár valóban történnek, de általában csak káros változásokat idéznek elő – a hasznos változások tudatos alkalmazkodási folyamatból erednek.
Egy példaként, mindez a „homoszexualitásért felelős gént” is új megvilágításba helyezi. Nyilvánvaló, hogy ez egy öngyilkos gén lenne, mivel akiben benne van, sokkal kisebb eséllyel fog szaporodni. Akkor hogy létezhet mégis? Úgy, hogy az újabb kutatások alapján lehetőség van arra is, hogy a környezeti hatások által újratermelődjön ez a gén, ha pedig a környezeti hatások nem aktiválják, oly mindegy, hogy ott van-e. A „genetikai determinizmus” tehát régen megbukott, de mindenhol ezt sulykolják: az anyák nagyobbik fele hajlamos lenne egy abortuszra csak azért, mert megállapítják, hogy a magzatban „homoszexualitásért felelős” vagy „homoszexualitást okozó” gén van. A gén nem „okoz” semmit, az indító impulzusok a környezetben vannak, a gén csak a reakció mikéntjét írja le.
V. Az evolúció mint hitrendszer
Az evolúció nem csak természettudományos munkahipotézis. Mivel a fent vázolt erőszakos és tisztességtelen terjesztési módszerek jelen vannak mind az iskolai tananyagban, mind a populáris kultúrában, az átlagember számára, aki nem szakértője az evolúciós biológiának, egy hitrendszer is: egy vezérlőideológia, tehát olyan ideológia, amely társadalmi relevanciája miatt jelentősen befolyásolja az emberek világnézetét és ezáltal hétköznapi viselkedését is.
Mit üzen nekünk a darwinizmus?
1. Az élet „véletlenül” jött létre, senki sem teremtette, nincs semmi oka, sem célja annak, hogy itt vagyunk, csak éppen így történt, s ha már így történt, valamit ki kéne találni, hogy mit kezdjünk magunkkal – vagy sem, végülis tök mindegy.
2. Az élet egyedüli objektív célja: az életben maradás, a túlélés. Ennek érdekében fő prioritásunk mindenkor a túlélési esélyünk növelése kell legyen. A biztonságra való törekvés fontosabb, mint pl. a boldogság keresése vagy a szabadság, stb. A biztonságra törekvés eszközei: a monotonitásra való törekvés, a nem várt események lehetőségének minimálisra csökkentése, szabályozással (erről lehet, hogy lesz egy külön bejegyzés), valamint a megfelelő stabil anyagi egzisztencia kiépítése, tartalékok felhalmozása, stb.
3. Az ember egy evolúciósan felkapaszkodott majom.
4. Az ember áll az evolúció csúcsán, s mivel az ember felett szellemi hatalom nincsen, ezért az ember írja a szabályokat önmagának
5. Az ember, mint egyedüli értelmes lény, képes szabályokat írni az „alatta lévő”, uralma alatt álló természetnek is.
Lehet összeesküvés-elméletet kiáltani, azonban nekem úgy tűnik, nagyszerű rabszolga lesz abból, aki az első 3 pontot elhiszi. A 4. pont adja a „tudományos” megalapozottságot a humanista ideológiának, s a ráépülő különböző ideológiai rendszereknek, különös tekintettel a liberalizmusra, amelyet az 1. pont is megtámogat (itt látható a liberalizmus és a nihilizmus közeli rokonsága is), valamint a szocializmus különböző fajtáira, amelyeket a 4. és 5. pont is megtámogat. Ezek nyitották meg az utat Marx „forradalmista” eszméihez, az Ausradieren gondolatához, stb. (Marx lelkes híve volt Darwinnak.) De gondoljunk csak bele: ha az ember egy majomból fejlődött ki Afrikában (3. pont), bizonyos embertípusok fejlettebbek másoknál, ergo egyes embertípusok közelebb állnak az állathoz! Nem véletlenül kezdték a nemzetiszocialista Németország iskoláiban ugyanolyan lelkesen tanítani az evolúciót, mint a Szovjetunióban a szocialista forradalom után. A fajelmélet teljes egészében a darwinizmusra épül. És ha vetünk egy pillantást az 5. pontra, hát elég szép ideológiai alapot ad a környezetpusztításra, a természet rendjének felforgatására, az embernek a természeten kívül-fölött való elhelyezésére, holott a 3. pont épp azt mondja, hogy az ember csak egy majom – de hát az ellentmondás ne zavarjon senkit. (Mindenesetre láthatjuk, hogy amikor a humanisták tűzik a környezetvédelmet a zászlajukra, a konfliktus feloldhatatlan.) Mint ahogy az 1. pont ellentmondása se zavarjon senkit: ha nincs se ok, se cél, akkor értelmetlené válik a „fejlődés” (evolúció) fogalma is.
Mindezeket figyelembe véve megállapíthatjuk, hogy a darwinizmus, legyen bármilyen ragyogó munkahipotézise a biológiának, egyúttal az egyik legkárosabb ideológia, amelyet valaha ember kitalált, s nagyobb szerepe volt a világ természetes rendjének szétverésében, mint bármely másik emberi gondolatnak a „felvilágosodás” történetében.
VI. Az emergencia
Az élet kezdetének, valamint a vertikális evolúciónak (lásd a VII. pontban) a problémájára válaszul felvetették az „emergencia”, mint új matematikai evolúciós csodaszer lehetőségét: az anyagban eleve benne foglaltatik az arra való hajlam, hogy fraktális formákat vegyen fel. Mint ahogy Darwin megfigyelte, hogy az egyik pinty rokonságban lehet a másikkal, s ebből kikövetkeztette, hogy a kenguruk is rokonságban vannak a pintyekkel, ez is hasonló mintát követ: látható, hogy hópehely létrejön magától, tehát csalán is fog. Ha el is fogadjuk, hogy az emergencia lehetősége jelentősen megnöveli az élet spontán létrejöttének esélyét, az ezzel kapcsolatos problémák:
1. Az emergencia elfogadása megdönti a bizonyosságot, hogy az élet egyetlen forrásból indult el, s így minden faj rokonságban van egymással.
2. Ha az emergencia igaz, az élet keletkezése laboratóriumban megismételhető. Hát, lássuk…
VII. Az evolúció matematikai cáfolatának lehetősége
A fajok közötti átalakulást általában két csoportra szokták osztani: „horizontális” és „vertikális” evolúció, vagy más kifejezéssel: mikro- és makroevolúció. Az előbbi mindössze ugyanazon élőlény-típusnak a kisebb átalakulásait jelenti, nagyrészben a meglevő genetikai információ torzításával, kisebb részben hasznos új géninformációk megjelenésével. Ez mérhető, megfigyelhető, bizonyítható tudományos tény, amely mellesleg nem cáfolja egyetlen történelmi vallásnak egyetlen tanítását sem. Az utóbbi viszont az egyes élőlény-típusok átalakulása, minőségi növekedés, új karakter és a hozzá tartozó, vaduonatúj leíró génszekvenciák megjelenése a semmiből – ez már csak spekuláció, nem kimutatható a gyakorlatban, s bár a tudósok szerint ez azért van, mert ez a folyamat csak évmilliók alatt történhet meg, valós paleontológiai bizonyíték sincs rá. Valószínűleg azért, mert soha nem történt meg.
A fajok átalakulása ugyanis fokozatosan történik, efelől nincs kétség: egyetlen csimpánz sem tud embergyereket szülni. A vertikális evolúcióban azonban olyan „evolúciós ugrások” vannak, amelyek fokozatos átalakulással nem valósíthatóak meg. Például:
1. a többsejtű élőlények úgy jöttek létre, hogy az egysejtűek kölcsönös előnyökért társultak egymással (ez eddig logikus), telepes élőlényeket létrehozva, majd a telepen belül az egyes sejtek specializálódni kezdtek a hatékonyabb kooperáció érdekében (ez szintén logikus), pl. a kívül lévő sejtek egy része megkeményedett, hogy kültakarót hozzon létre, mások magukra kellett, hogy vállalják a teljes telep „etetését”, hiszen a középen lévők legfeljebb a szomszédaikat ehették volna, az meg nem valami jó, stb. Van az elméletnek egy szépséghibája: a többsejtű organizmusokban a sejtek DNS-e az egész organizmust írja le, nem csak az adott sejtet. Tessék mondani, ez hogy történt fokozatosan?
2. A kígyóknak állítólag kezdetleges lábacskáik vannak, amelyek évmilliók után járásra is alkalmasak lesznek majd. Egyrészt ezek a „kezdetleges lábacskák” már most is használatban vannak szaporodáskor, másrészt ha nőni kezdenek, egyre inkább csak útban lesznek majd a kígyóknak, s csak sok-sok generáció után tud elkezdeni mászni velük – addig csak akadályozzák a siklásban.
3. Az őshüllő elülső végtagjai állítólag szárnyakká alakultak. A probléma ugyanaz: lesz sok-sok ezer olyan generáció, amelyek járni már nem tudnak vele, repülni még nem.
Végtelenségig lehetne példákat sorolni ugyanerre a kaptafára. A vertikális (vagy makro-) evolúciót nem azért nem rekonstruálják laboratóriumi állatokkal, mert „túl sokáig tartana”. Egyrészt elkezdeni azért el lehetne, nem? Másrészt ha már tudjuk, mi a kívánt végeredmény (lábakon mászó kígyó, azaz immár gyíkféle), irányított szelekcióval a folyamat rendkívül felgyorsítható lenne. Lenne, ha egyáltalán lehetséges lenne. Valószínűleg voltak ilyen irányú kísérletek, csak a fenti probléma miatt kudarcba fulladtak.
A probléma matematikai megközelítése:
A fokozatos átalakulás problémája, hogy ha az átalakulási folyamat elején „bemenő” élőlény túlélési esélyénél magasabb eséllyel bír a folyamat végén „kijövő” élőlény, a köztes fázisok túlélési esélye általában nem lineárisan növekszik, sőt sokszor egyáltalán nem monoton növekvő. Ehelyett létrejön az általam „hasznossági szakadéknak” nevezett képződmény:
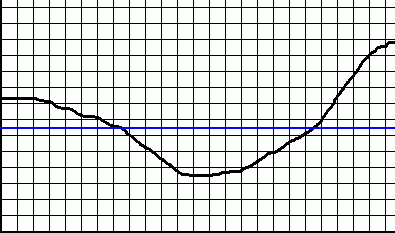
A vízszintes tengelyen az idő múlása és a generációk egymásra következése kap helyet, a függőlegesen pedig a „hasznosság”, szigorúan túlélési és utódnemzési esélyként kifejezve. Ez természetesen egy véletlenszerű görbe a probléma nagyjábóli érzékeltetésére, nem egzakt számértékeket mutat, és minden esetben más a görbe, a minimumhelyig sem feltétlenül monoton csökkenő, azután sem feltétlenül monoton növekvő, stb.
Egy „rosszabb” mutáció, mint látható is, egy határig életben maradhat. Egy rövidebb lábú antilopnak kisebb az esélye, hogy elmeneküljön a gepárd elől, de ez nem jelenti azt, hogy szükségszerűen kudarcra van ítélve. A kék vonal mutatja azt a szintet, ahol az adott mutációs szekvencia kihalásra, illetve a saját elődje általi kiszorításra van ítélve. Az átalakulás ezen a ponton megszakad – a makroevolúció matematikai képtelenség.
A hernyó sem versenyképes a természetben a lepkévé változás ideje alatt, elrejtőzik és begubózik, a viszonylag rövid átalakulási folyamatot így vészeli át. Azonban egy évmilliókig tartó átalakulás idejére a bálnává változó tehén, miközben éppen úszóleckéket vesz, nem írhatja ki egy táblára: „evolúció alatt – légyszi ne egyél meg”. Nem kerülheti el az élet színpadán való megmérettetést több generáción át, s így nem kerülheti el a kihalást sem. A darwinizmus feltételezi, hogy az összes átalakulási fázis elégséges versenyképességgel rendelkezik.
Ez egyúttal megmagyarázza azt is, hogy a kisebb átalakulások miért lehetségesek: a hasznossági függvényük akár teljesen lineáris is lehet, de mindenesetre szigorúan monoton növekvő, lásd a zsiráf nyakának esetét, ahol a hosszabb nyak előnyösebb, nincs hátrányos köztes fázis.
Természetesen egy evolúciós folyamat kezdete és vége között több különböző lehetséges átmeneti útvonal van, s mindegyiknek más a hasznossági függvénye. Így meg kell találnunk azt az útvonalat, ahol egyetlen érték sem megy a kék vonal alá (amit persze szintén meg kellene határoznunk, hogy hol van!), s megoldottuk a problémát! Ha így nézzük, tulajdonképpen az egyes lehetséges átalakulási fázisokat, az átalakulóban lévő tulajdonság pillanatnyi lehetséges állapotait egy multidimenzionális gráf pontjaiként ábrázolhatjuk, ahol természetesen minden gráfpontnak sok közvetlen szomszédja van, mint mutációs lehetőségek. Az egyszerűség kedvéért ezt képzeljük el egy kétdimenziós gráfként, ahol egyik pontból a másikba szeretnénk eljutni, s erre rengeteg különöböző bejárási útvonalunk van. A 3. dimenziót pedig használjuk fel arra, hogy adjunk minden gráfpontnak egy „magasságot”, Z koordinátát, amely a túlélési esélyét mutatja. Így hasznossági hegyeket-völgyeket fogunk kapni. A kék vonal kiszámolása – valószínűleg Nash-egyensúly által – a legnagyobb problémánk, de ha sikerült, az egydimenziós vonalból egy kétdimenziós síkfelület lesz, így ezt felfoghatjuk „vízszintnek is”, mintha a völgyeink víz alá kerülnének, s mivel a kék vonal mutatta az életképesség határát, csak azok az állapotok életképesek, amelyek a vízszint felett vannak. Ez egyúttal azt is jelenti, hogy a hasznossági hegyekből hasznossági szigetek lesznek, s az összes ma élő fajt ezen szigeteken fogjuk megtalálni, túlnyomó részüket a középen levő csúcsok táján. Az egyes szigeteket vékony, kacskaringós „hidacskák” kötik össze, amelyek az átalakulás lehetőségét bizonyítják. DE! Amennyiben nincs az összes sziget között átjárhatóság – s meggyőződésem, hogy nincs – hanem szigetcsoportokat alkotnak, s az egyes szigetcsoportok között NEM LÉTEZIK átjárási útvonal, minden lehetséges útvonal víz alatti pontokon keresztül vezet, akkor matematikailag bizonyítottuk, hogy a Földön élő összes létformának NEM LEHET közös őse, az egyes szigetcsoportok között nincs evolúciós átjárhatóság, még irányított mutációval sem, így szeparált tudatos teremtés eredményei – amennyiben egy adott szigetcsoporton található legegyszerűbb formula már annyira komplex, hogy emergenciával sem, sem semmilyen egyéb módon nem magyarázható meg a spontán létrejötte. Ez valószínűleg így lesz, hiszen a fenti példa alapján az egysejtűek eleve külön szigetcsoporton eveznek, így bizonyíthatóan nem ősei a multicelluláris lényeknek.
Ezért kerestem én matematikust a rádióban, nem pedig azért, hogy mindezekre rájöjjön helyettem. Nekem egy olyan matematikus kellene, aki mindezt a számok szintén le tudja írni, s így együtt be tudjuk bizonyítani, hogy a Földön létező egyes élőlény-típusok: tudatos tervezés eredményei.
Adalék: a vertikális evolúció további problémája
Az evolúció állítólagos hajtóereje, hogy az élőlények az átalakulás által növelik a túlélési esélyeiket. A probléma az, hogy a vertikális evolúció általában hosszú távon is csökkenti a túlélési esélyeket. Szemben a fenti görbével, a végpont általában alacsonyabban lesz, mint a kezdőpont. Ha Budapestre ma este atombombát dobnak, miből mennyi marad ezek közül: moha, légy, patkány, ember? Látható, hogy egy élőlény minél bonyolultabb, annál „kényesebb”, annál könnyebben tönkremegy. Akkor mi hajtja a makro-evolúciót?
*Helyesbítés: Az nem simaizomszövet, valóban a bélcsatornával van összekötve (székletürítéskor használatos), de mivel szomatikusan vezérelhető és nagy erőkifejtésre képes izomzat, ez alapján harántcsíkolt kell hogy legyen, és félig-meddig a vázizomzathoz sorolható, gyakorlatilag határeset. („Hivatalosan” biztos, hogy ide vagy oda van sorolva, nem tudom.) Ettől függetlenül a farokcsont használatban van, tehát a leírtak lényege nem változik. A 2012 Blog írásához kurva sok féle szakterületbe kell belefolynom, nincs ember, aki ezekhez mind értene olyan szinten, hogy ne vétene néha a tényszerűség ellen.